1) Il metabolismo e' regolato localmente, nelle singole cellule, in funzione del fabbisogno energetico (che la cellula stima dal rapporto tra AMP e ATP) e centralmente, mediante ormoni specifici.
2) La regolazione locale (intracellulare) e' principalmente governata dalla Proteina Kinasi-AMP dipendente (AMPK). L'AMPK e' un enzima trimerico altamente conservato negli eucarioti, formato dalle tre subunita' α, β e γ. La subunita' α possiede una treonina (Thr 172) che puo' essere fosforilata; in questo stato si attiva ed e' capace di fosforilare a sua volta varie proteine bersaglio. L'attivazione della subunita' α avviene grazie ad un cambiamento conformazionale che consegue al legame dell'AMP sulla subunita' γ ed espone la Thr 172. Il legame dell'ATP alla stessa subunita' ha invece un effetto opposto e stabilizza la conformazione quaternaria inattiva, nella quale il residuo Thr 172 e' nascosto.
3) I principali bersagli dell'AMPK sono:
a) la fosfofruttokinasi 2 cardiaca e di altri tessuti che converte fruttosio-6-fosfato (F6P) in fruttosio-2,6-bisfosfato (F2,6P2); quest'ultimo attiva la fosfofruttokinasi che converte F6P in F1,6P2 e attiva quindi la glicolisi.
b) Negli epatociti la fosforilazione mediata dall'AMPK inibisce vari enzimi che catalizzano reazioni anaboliche:
L'acetil-CoA carbossilasi (ACC) epatica, che viene inibita, bloccando la biosintesi degli acidi grassi.
La β-idrossi-β-metil glutaril-CoA reduttasi epatica, che viene inibita, bloccando la biosintesi del colesterolo.
La glicogeno sintetasi epatica, che viene inibita, bloccando la biosintesi del glicogeno
c) Nel muscolo scheletrico il calo della concentrazione del malonil-CoA dovuto alla fosforilazione dell'ACC del miocita promuove la β-ossidazione degli acidi grassi. Inoltre promuove l'esposizione dei trasportatori del glucosio GLUT4, favorendo l'assorbimento del glucosio.
d) Nel tessuto adiposo viene fosforilata la lipasi, con effetto inibitorio.
4) La regolazione ormonale del metabolismo e' molto complessa; vi partecipano:
a) insulina e glucagone: sono prodotti nelle isole di Langerhans del pancreas e controllano la glicemia e il rapporto tra metabolismo di zuccheri e grassi
b) adrenalina e noradrenalina: sono prodotte dalla midolla surrenale e contribuiscono al controllo del metabolismo glicidico
c) resistina, adiponectina e altri: sono prodotti dal tessuto adiposo e regolano la sensibilita' dei diversi tipi cellulari all'insulina
d) glicocorticoidi: sono prodotti dalla corteccia surrenale e regolano l'espressione genica
e) leptina, grelina e altri: controllano la sensazione di fame e la regolazione dell'apporto calorico
f) tiroxina e triiodotironina: sono prodotti dalla tiroide e controllano il disaccoppiamento della fosforilazione ossidativa e, conseguentemente, il metabolismo basale.
5) L'insulina e' unormone proteico ed e' il piu' importante regolatore del metabolismo glicidico e lipidico; il diabete di tipo I, nel quale le cellule β delle isole di Langerhans muoiono per un attacco autoimmune e' una malattia rapidamente mortale, se non trattata con la somministrazione di insulina esogena. Nella maggioranza delle cellule, l'insulina promuove il metabolismo glicidico, a discapito di quello lipidico. La principale (ma non unica) azione dell'insulina e' quella di legarsi ad un recettore di membrana la cui attivazione promuove l'esposizione sulla membrana stessa del trasportatore GLUT4 (altrimenti presente in forma di vescicole citoplasmatiche) che permette al glucosio di entrare nella cellula.
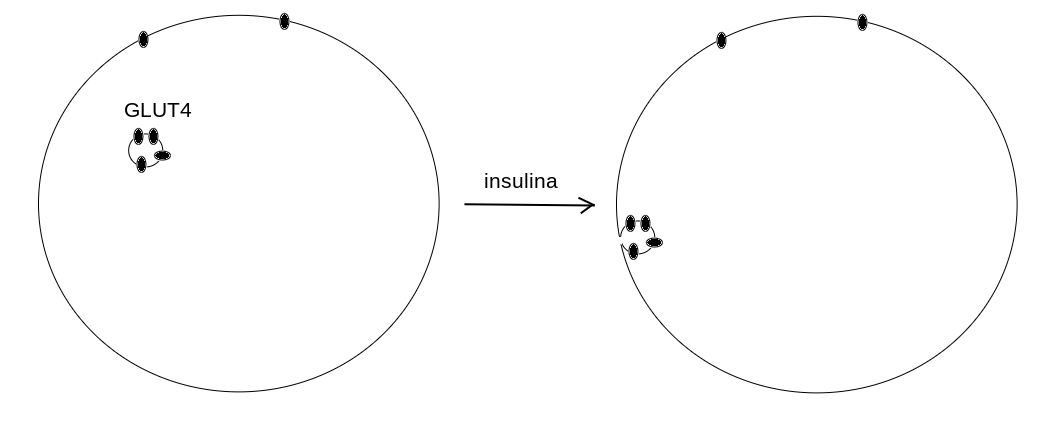
6) La carenza di insulina nel diabete mellito di tipo I fa si' che il metabolismo cellulare si rivolga soprattutto all'utilizzazione dei grassi anziche' del glucosio, e in parte a quella degli aminoacidi. Questo cambiamento qualitativo del metabolismo crea pero' un grave problema, la deplezione dell'ossalacetato nel mitocondrio, dovuta alle reazioni cataplerotiche che sottraggono metaboliti intermedi al ciclo di Krebs. Infatti nei mammiferi la principale reazione anaplerotica e' la carbossilazione del piruvato operata dalla piruvato carbossilasi, che converte questo metabolita in ossalacetato; e la principale sorgente di piruvato e' la glicolisi. La β-ossidazione degli acidi grassi produce quindi piu' acetil-CoA di quanto ne possa essere utilizzato nel ciclo di Krebs e stimola la chetogenesi. I corpi chetonici prodotti in eccesso vengono rilasciati nel sangue causando una grave chetoacidosi metabolica, che puo' provocare il coma. Il paziente in coma chetoacidosico va incontro a una riduzione della stimolazione dei centri respiratori che vanifica il compenso respiratorio dell'acidosi e se non trattata porta al decesso.
7) Il glucagone e' prodotto dalle cellule α delle isole di Langerhans del pancreas e svolge una funzione opposta a quella dell'insulina. Il glucagone viene secreto in risposta all'ipoglicemia e promuove il rilascio di glucosio dal fegato (glicogenolisi).
| organo | insulina | glucagone |
| sangue | diminuzione della glicemia | aumento della glicemia |
| muscolo | assorbimento del glucosio glicogenosintesi | nessun effetto |
| tessuto adiposo | assorbimento del glucosio lipogenesi riduzione della lipolisi | aumento della lipolisi |
| fegato | assorbimento del glucosio glicogenosintesi lipogenesi riduzione della gliconeogenesi | gliconeogenesi glicogenolisi |
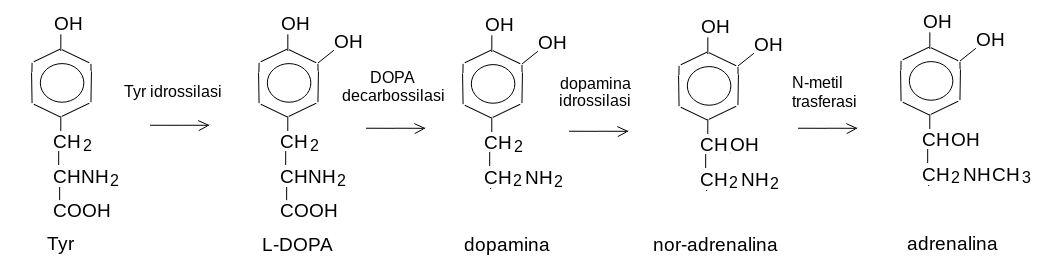
8) Adrenalina e nor-adrenalina stimolano il rilascio di glucagone e provocano nella cellula effetti simili a quelli del glucagone. Sono composti a basso peso molecolare prodotti dalla midolla surrenale e dal tessuto nervoso a partire dall'aminoacido tirosina (che a sua volta puo' essere prodotto dalla fenilalanina) ed hanno funzioni sia di ormoni che di neuromediatori (come anche il loro precursore dopamina). La via biosintetica e' come segue:
9) I principali glicocorticoidi sono il cortisolo e il cortisone, prodotti dalla corteccia surrenale a partire dal colesterolo. Attraversano la membrana cellulare e si legano ad un recettore nucleare che agisce come promotore dell'espressione genica.
10) Il tessuto adiposo produce vari ormoni di natura proteica che antagonizzano con l'insulina, riducendo l'espressione del suo recettore. I principali sono la resistina e l'adiponectina. Questi ormoni possono contribuire a causare iperglicemia e diabete mellito di tipo II (insulino-resistente, periferico).
| resistina, adiponectina | aumento della glicemia insulino-resistenza |
| leptina | soppressione della sensazione di fame |
11) La regolazione dell'apporto calorico dipende anch'essa da ormoni. L'ipotalamo secerne una famiglia di peptidi (peptidi Y) che stimolano la sensazione neurologica della fame e inducono l'animale a mangiare. La secrezione dei peptidi Y e' inibita dall'ormone leptina una proteina codificata dal gene Ob (per obesita') espressa e secreta nel sangue dal tessuto adiposo. Quanto piu' l'animale (o la persona) possiede tessuto adiposo tanto piu' produce leptina e tanto piu' inibisce la produzione ipotalamica di peptidi Y e quindi lo stimolo a mangiare; in pratica la leptina informa il sistema nervoso centrale sull'entita' delle riserve di grasso nel tessuto adiposo. La leptina (col gene Ob) fu scoperta in studi su ceppi di topi geneticamente obesi a causa di mutazioni del gene Ob.
La grelina e' un ormone proteico prodotto dalla mucosa gastrica che promuove la sensazione di fame. E' prodotta quando lo stomaco e' vuoto, mentre la produzione e' inibita dalla distensione gastrica; in pratica segnala all'ipotalamo lo stato di distensione dello stomaco.
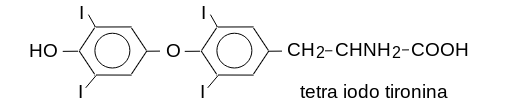
12) Gli ormoni tiroidei: tiroxina (tetraiodo tironina, T4) e triiodotironina (T3) sono dei derivati iodinati dell'aminoacido tirosina (la tiroide e' il principale organo che utilizza e richiede lo iodio). Hanno vari effetti sul metabolismo, il principale dei quali e' il disaccoppiamento della fosforilazione ossidativa (termogenesi, aumento del consumo calorico). Sono conservati nella ghiandola sotto forma di aminoacidi modificati posttraduzionalmente nella proteina tireoglobulina, che deve essere digerita per permettere il rilascio dell'ormone nel sangue.