1) Il ciclo di Krebs e' una via metabolica interamente mitocondriale di fondamentale importanza per la produzione di energia. Converte acetil-CoA in due molecole di CO2. L'equazione complessiva del ciclo e' la seguente:
CH3-CO-SCoA + FAD + 3 NAD+ + GDP + Pi --> 2 CO2 + CoASH + FADH2 + 3 NADH + GTP
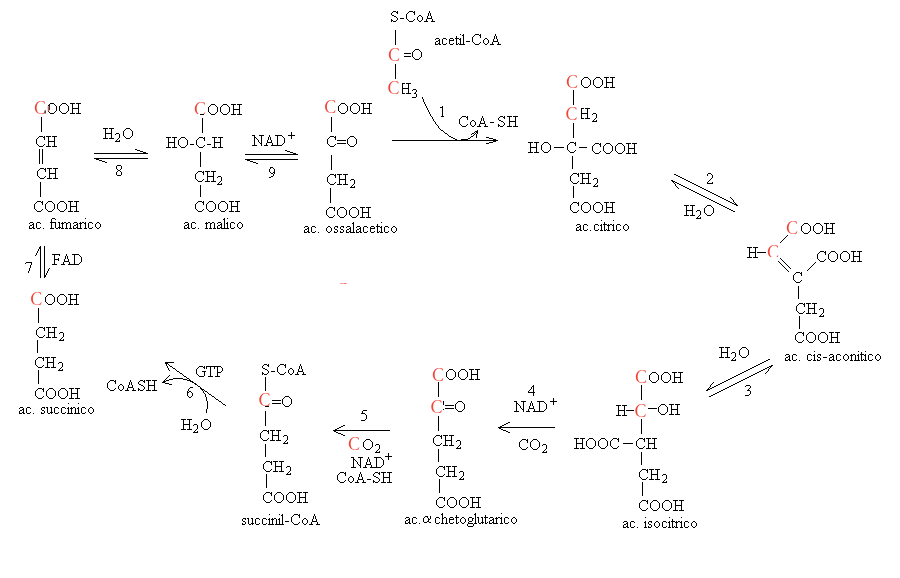
2) Le reazioni del ciclo sono le seguenti:
Sono evidenziati in rosso gli atomi di carbonio dell'acetil-CoA e si nota che in ogni ciclo viene eliminata una molecola di CO2 proveniente dall'acido ossalacetico e una dall'acetile.
3) Gli enzimi del ciclo
Il primo enzima del ciclo e' la citrato sintetasi che catalizza la sintesi di citril-CoA a partire da acetil-CoA e acido ossalacetico (reazione 1). Il citril-CoA e' instabile e decade spontaneamente e molto rapidamente a citrato e CoA-SH, quindi non viene considerato tra gli intermedi del ciclo. La formazione del citrato e' essenzialmente irreversibile (Keq > 105).
L'aconitasi catalizza due reazioni del ciclo: l'estrazione e l'inserimento di una molecola d'acqua dal citrato e la sua inversa, l'addizione di una molecola d'acqua all'aconitato (reazioni 2 e 3). Entrambe le reazioni sono reversibili e le tre molecole citrato, aconitato e D-isocitrato sono tra loro in equilibrio; pero' poiche' soltanto l'isocitrato viene consumato nella successiva reazione del ciclo, l'equilibrio viene drenato verso quest'ultimo.
L'isocitrato deidrogenasi (o decarbossilasi) ossida e decarbossila l'isocitrato producendo una molecola di CO2 e una di acido α-chetoglutarico e riducendo al contempo una molecola di NAD+ (reazione 4).
L'α-chetoglutarato deidrogenasi (o decarbossilasi) ossida e decarbossila l'α chetoglutarato e coniuga il prodotto col CoA. In questa reazione viene prodotta la seconda molecola di CO2 e viene ridotta una second molecola di NAD+ (reazione 5). Il meccanismo di azione di questo enzima (decarbossilazione ossidativa dell'α chetoacido) e' analogo a quello della piruvato deidrogenasi.
L'idrolisi del tioestere e il rilascio del CoA-SH e' catalizzato dalla succinil-CoA sintetasi che catalizza anche la fosforilazione di una molecola di GDP (equivalente ad un ADP; reazione 6).
La succinato deidrogenasi ossida il succinato a fumarato e trasferisce due elettroni su un FAD interno all'enzima (reazione 7). Questo enzima e' molto peculiare perche' e' associato alla membrana mitocondriale interna e fa parte tanto del ciclo di Krebs quanto della catena respiratoria mitocondriale; infatti trasferisce la coppia di elettroni dal FADH2 al coenzima Q.
La fumarasi addiziona H2O sul doppio legame dell'acido fumarico, convertendolo in acido malico (reazione 8).
La malato deidrogenasi ossida l'acido malico a ossalacetico, con riduzione di una molecola di NAD+ (reazione 9) e chiude il ciclo.
4) Gli atomi di carbonio eliminati sotto forma di CO2 derivano dal C1 dell'acido ossalacetico e dal C1 dell'acetile legato al Coenzima A:
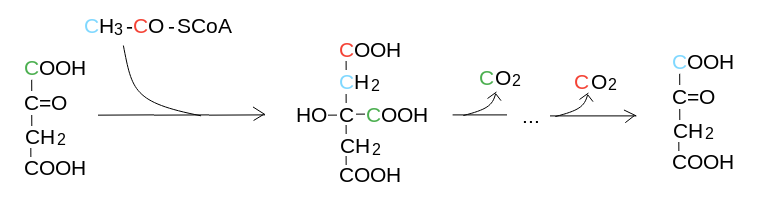
5) Cataplerosi. Oltre a produrre energia, il ciclo produce metaboliti intermedi che possono essere utilizzati da altre vie metaboliche. Le reazioni che sottraggono intermedi al ciclo di Krebs sono chiamate reazioni cataplerotiche (dal greco: che svuotano il ciclo). Le principali reazioni cataplerotiche sono le seguenti:
a) il citrato puo' essere trasferito al citoplasma cellulare dove viene riconvertito in acetil-CoA (per la biosintesi di acidi grassi e colesterolo) e ossalacetato, che puo' essere utilizzato per la gliconeogenesi, dall'enzima P-enolpiruvato carbossichinasi, oppure restituito al mitocondrio.
b) L'α-chetoglutarato viene utilizzato dalle transaminasi per la biosintesi dell'acido glutamico e per la transaminazione degli aminoacidi.
c) Il succinil-CoA viene utilizzato per la biosintesi dell'acido δ-aminolevulinico, primo metabolita della biosintesi delle porfirine.
d) Il malato viene utilizzato per la gliconeogenesi
e) L'ossalacetato viene utilizzato per la biosintesi dell'acido aspartico.
6) Anaplerosi. Poiche' la via e' un ciclo del quale le reazioni cataplerotiche impediscono la chiusura e quindi in ultima analisi sottraggono ossalacetato, e' necessario che gli intermedi del ciclo vengano riforniti, da reazioni anaplerotiche (dal greco: che riempiono il ciclo). Queste sono:
a) la produzione di ossalacetato a partire dal piruvato, catalizzata dalla piruvato carbossilasi mitocondriale. Questa e' la piu' importante reazione anaplerotica e la principale fonte di ossalacetato nel mitocondrio:
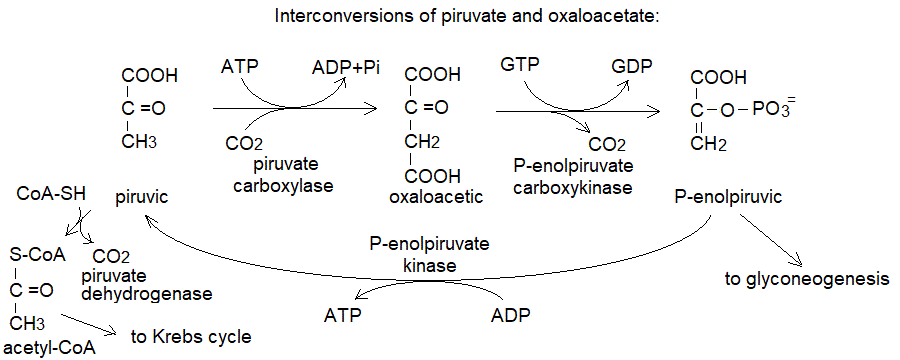
b) La produzione di ossalacetato a partire dall'acido aspartico (o asparagina), catalizzata dall'aspartico transaminasi.
c) La produzione di α-chetoglutarato a partire dall'acido glutamico (o glutamina).
d) La produzione di succinil-CoA, prodotto finale del metabolismo del propionil-CoA, prodotto dalla β-ossidazione degli acidi grassi a numero dispari di C, o dal metabolismo degli aminoacidi Met, Val, Ile.
e) La produzione di fumarato derivante dal metabolismo degli aminoacidi Phe e Tyr.
7) I fenomeni anaplerotici e cataplerotici sono molto importanti per ovvie ragioni fisiologiche. In medicina hanno conseguenze peculiari in molte malattie genetiche, perche' fanno si che il difetto genetico di una via metabolica possa ripercuotersi su altre vie metaboliche, apparentemente non correlate, ma in realta' collegate alla prima da reazioni anaplerotiche o cataplerotiche: un esempio classico e' la chetoacidosi diabetica, che deriva dall'accumulo di metaboliti degli acidi grassi in pazienti che non metabolizzano correttamente il glucosio.