I concetti di etnia, popolazione e razza sono piuttosto ambigui; pero' hanno importanza per la medicina e non possono essere ignorati.
- Una popolazione e' un gruppo che vive in un ambiente comune e condivide linguaggio e cultura; puo' o meno presentare frequenze geniche diverse da quelle delle popolazioni confinanti.
- La razza e' invece definita su base genetica ed indica un gruppo, interno ad una specie, che ha caratteristiche genetiche comuni, ben definite. In pratica una razza puo' esistere soltanto se i membri della specie per qualche ragione geografica o culturale praticano una rigorosa endogamia e si sposano soltanto all'interno del gruppo di riferimento. In linea di massima la gran parte delle razze animali e vegetali sono artificiali e prodotte dall'uomo mediante il controllo degli incroci. Lasciata a se' la razza si disperde e mescola con gli altri membri della specie. Tutti i genetisti moderni concordano sul fatto che la specie umana non puo' essere classificata in razze distinte: non esistono vere razze umane; cio' che si osserva sono gradienti di frequenze geniche tali per cui una popolazione umana puo' essere geneticamente distinguibile da un'altra, ma la variabilita' interna alla popolazione risulta sempre molto grande rispetto alla variabilita' genetica che distingue le due popolazioni.
- L'etnia e' un concetto intermedio tra i due precedenti che puo' essere di volta in volta utilizzato per indicare gruppi genetici, gruppi culturali o gruppi individuati da entrambe le caratteristiche.
Per chiarire il concetto di razza e' utile considerare un esempio molto semplificato che presento qui sotto. La semplificazione e' necessaria sia per evitare una trattazione matematica molto difficile, sia per facilitare la logica del ragionamento.
Immaginiamo una popolazione animale di 2.000 individui appartenenti tutti alla medesima specie, ma suddivisa in due sottopopolazioni distinte, che chiameremo P1 e P2, ciascuna composta da 1.000 individui. I membri di queste due popolazioni, che in teoria sono interfecondi, sono rigorosamente endogamici: i membri di P1 si accoppiano soltanto con altri membri di P1 quelli di P2 fanno lo stesso. Immaginiamo che i membri di P1 e P2 siano distinguibili per un qualche carattere; ad esempio gli uni cacciano di giorno, gli altri di notte.
Supponiamo ora di fare sequenziare un solo gene nel genoma di questi animali e supponiamo che questo gene presenti due alleli, che chiamiamo A e a. I genotipi possibili sono AA, Aa e aa, e obbediscono alla legge di Hardy-Weinberg; non c'e' selezione nei confronti dell'uno o dell'altro. Se tra le due sottopopolazioni esiste una differenza genetica questa si manifesta nelle frequenze geniche; supponiamo, ad esempio che la situazione che la nostra analisi genica ci rivela sia quella riportata nella Tabella qui sotto:
| sottopopolazione 1 | sottopopolazione 2 | popolazione totale | |
| numerosita' | 1.000 | 1.000 | 2.000 |
| frequenza dell'allele A | 800 / 1.000 = 0,8 | 200 / 1.000 = 0,2 | 1.000 / 2.000 = 0,5 |
| frequenza dell'allele a | 200 / 1.000 = 0,2 | 800 / 1.000 = 0,8 | 1.000 / 2.000 = 0,5 |
| frequenza del genotipo (AA) | 0,64 | 0,04 | 0,34 (atteso 0,25) |
| frequenza del genotipo (Aa) | 0,32 | 0,32 | 0,32 (atteso 0,5) |
| frequenza del genotipo (aa) | 0,04 | 0,64 | 0,34 (atteso 0,25) |
Il sistema descritto presenta una distribuzione binaria (questo non sarebbe vero se esistessero piu' di due sole varianti alleliche, come quasi sempre accade; le distribuzioni non binarie richiedono una statistica molto piu' complessa di quella presentata in questo esempio). I parametri statistici di un sistema binario sono facilissimi da calcolare: e' sufficiente assegnare i valori di 0 e 1 alle due variabii (ad es. A=1, a=0) e calcolare medie e varianze con le formule apposite. La tabella puo' quindi essere estesa in questo modo:
| sottopopolazione 1 | sottopopolazione 2 | popolazione totale | |
| numerosita' | 1.000 | 1.000 | 2.000 |
| frequenza dell'allele A | 800 / 1.000 = 0,8 | 200 / 1.000 = 0,2 | 1.000 / 2.000 = 0,5 |
| frequenza dell'allele a | 200 / 1.000 = 0,2 | 800 / 1.000 = 0,8 | 1.000 / 2.000 = 0,5 |
| "valore" della media (A=1, a=0) | 0,8 | 0,2 | 0,5 |
| varianza σ2 = freq.(A) x freq.(a) | 0,16 | 0,16 | 0,25 |
| devianza = N x σ2 | 160 | 160 | 500 |
Si chiama devianza totale (Dt) la devianza misurata nella popolazione totale; devianza within la devianza interna a ciascuna popolazione (Dw = D1, D2); e devianza between (Db) la devianza tra le medie delle due popolazioni. Vale la regola: Dt = Dw + Db. Questi parametri sono riportati nella seguente tabella:
| Dt | Dw = D1 + D2 | Db | Dw+Db |
| 500 | 160 + 160 = 320 | 1.000x(0,8-0,5)2 + 1.000x(0,2-0,5)2 = 180 | 320+180 = 500 |
Le differenze statistiche nei pool genici delle due sottopopolazioni sono state calcolate. Non vi sono dubbi sul fatto che le due sottopopolazioni sono geneticamente distinte. La domanda e': sono razze; gruppi; popolazioni; etnie? Evidentemente la distinzione tra questi concetti e' arbitraria e si basa su convenzioni. C'e' un generale accordo tra genetisti delle popolazioni sul definire razza un gruppo che differisca geneticamente dalla specie di appartenenza per una devianza between pari o superiore al 25% della devianza totale della specie. Nel caso considerato in questo esempio Db / Dt = 180 / 500 = 0,36 quindi secondo questa definizione P1 e P2 sono razze distinte della medesima specie. Si noti che i genetisti delle popolazioni calcolano questi parametri su un gran numero di geni, di ciascuno dei quali sono in genere presente parecchi alleli. Per questo tipo di calcoli sono richieste formule molto piu' complesse di quelle applicabili alle distribuzioni binarie come quella considerata.
Nelle popolazioni umane Db e' sempre molto piccolo rispetto a Dw e Dt; quindi la specie umana non puo' essere considerata come suddivisa in razze. Pero', se si isolano due popolazioni geograficamente e culturalmente distanti e si ignorano le popolazioni geograficamente e culturalmente interposte (ad es. europei e cinesi, ignorando le popolazioni del vicino e medio oriente), si possono trovare valori di Db relativamente grandi e avere l'erronea impressione che esistano razze umane. Questa impressione viene corretta quando si considera che europei e cinesi non vanno confrontati isolatamente tra loro, ma nel contesto dell'intera umanita': Dt non va riferito alla somma artificiale Europei+Cinesi ma all'umanita'. I genetisti di popolazione che studiano le popolazioni umane sono sostanzialmente concordi nel sostenere che le distribuzioni geniche della specie umana seguono gradienti geografici e culturali, piuttosto che raggrupparsi in razze. Si noti che i caratteri descritti appartengono alle popolazioni, non agli individui: la sottopopolazione P1 del nostro esempio, contiene individui aa, sebbene rari, geneticamente identici agli individui aa frequenti nella sottopopolazione 2. Le frequenze geniche sono caratteristiche dei gruppi, non possedute individualmente da nessuno dei membri del gruppo.
Qual e' l'importanza dello studio genetico delle popolazioni umane? Popolazioni geneticamente distanti tra loro possono presentare grandi variazioni nell'incidenza o nella prevalenza della stessa malattia, o diverse risposte agli stessi farmaci. La cosiddetta medicina personalizzata e' in realta' una medicina applicata a gruppi geneticamente omogenei (o almeno omogenei rispetto ai caratteri genetici ritenuti di volta in volta rilevanti).
Si consideri il seguente esempio: nell'isola abitata dalle due sottopopolazione P1 e P2 si sparge una epidemia. L'attack rate dell'epidemia e' indipendente dalla costituzione genetica degli individui ed e' del 90%; la letalita' della malattia e' invece dipendente dalla costituzione genica degli individui e risulta dello 0,1 % per il genotipo AA e del 10% per i genotipi che contengono il gene a (Aa e aa). Ad esempio si puo' immaginare che il gene codifica per una proteina di membrana (P), che costituisce il recettore del virus. La variante codificata dall'allele A (PA) ha bassa o bassissima affinita' per il virus, quella codificata dal gene a ha alta affinita' per il virus (Pa). Sulla membrana cellulare, un individuo AA espone PA; un individuo Aa espone sia PA che Pa, mentre un individuo aa espone la sola Pa. Se sulla membrana cellulare e' presente Pa la cellula e' facilmente infettata dal virus. La mortalita' osservata nelle due sottopopolazioni e nella popolazione totale, dopo che l'epidemia ha compiuto il suo corso e' la seguente:
| sottopopolazione 1 | sottopopolazione 2 | popolazione totale | |
| casi di malattia tra gli individui AA | 576 | 36 | 612 |
| decessi tra gli individui AA | 6 | 0 | 6 |
| casi di malattia tra gli individui Aa e aa | 324 | 864 | 1.188 |
| decessi tra gli individui Aa e aa | 32 | 86 | 118 |
| casi di malattia | 900 | 900 | 1.800 |
| decessi | 38 | 86 | 124 |
Un esempio reale: la variabilita' genetica della fosfatasi alcalina placentare umana.
Della fosfatasi alcalina prodotta dalla placenta umana esistono tre varianti alleliche maggiori, piu' un certo numero di varianti rare. La frequenza di queste varianti, indicate per la diversa velocita' di migrazione nell'elettroforesi come S (slow), I (intermediate) e F (fast), in diverse popolazioni umane e' riportata nella Tabella qui sotto (da uno studio classico di Roychoudhury e Nei del 1988):
| Regione | S | F | I | altre |
| Inghilterra | 0,637 | 0,270 | 0,085 | 0,008 |
| Italia | 0,661 | 0,256 | 0,075 | 0,007 |
| India occidentale | 0,701 | 0,217 | 0,066 | 0,016 |
| Tailandia | 0,746 | 0,081 | 0,165 | 0,008 |
| Giappone | 0,724 | 0,038 | 0,236 | 0,003 |
| Nigeria | 0,942 | 0,019 | 0,039 | - |
| Inuit del Canada | 0,556 | 0,142 | 0,296 | 0,006 |
| Papua della Nuova Guinea | 0,880 | 0,050 | 0,068 | 0,002 |
Considerazioni storiche.
La storia del concetto di razza e' peculiare perche' sebbene le diverse popolazioni fossero note fin dalla piu' remota antichita' e classificate variamente in base a lingua, religione, tratti fisici, etc., la formalizzazione "moderna" del concetto avvenne soltanto nel XIX secolo con i trattati di De Gobineau (Essai sur l'inegalite' des races humaines 1853), Knox (The races of man, 1850) e Grant (The Passing of the Great Race: Or, The Racial Basis of European History, 1916). Questi autori scrivevano in un'epoca coloniale, nella quale l'incontro con popolazioni diverse era causato dalla conquista e sottomissione di popoli e intendevano in qualche modo spiegare e giustificare la supremazia occidentale. La premessa generale di questa teorizzazione era che le razze non solo esistessero, ma che fossero una caratteristica filogenetica dell'evoluzione umana; che cioe' l'umanita' fosse, fin dalle sue origini, polifiletica, e costituita da gruppi distinti. Il mantenimento della "purezza" razziale originaria era quindi alternativo al "meticciato", ritenuto fonte di indebolimento della razza. L'idea di per se' era assurda, ma parzialmente giustificata dalle scarse conoscenze di biochimica e genetica dell'epoca. Infatti e' impossibile che due gruppi animali distinti evolvano in modo tale da diventare interfecondi, come ovviamente sono le diverse popolazioni umane. La speciazione avviene in un gruppo piu' o meno segregato, e piu' o meno grande di individui, e la differenziazione in razze, se avviene, diversifica un gruppo inizialmente omogeneo.
Perche' gli antichi non videro le razze? In epoca classica (greco-romana, ma anche araba) e fino al Rinascimento non esisteva il concetto di razze umane come inteso in epoche successive; eppure gli antichi erano grandi viaggiatori e conquistatori e conoscevano popoli molto diversi tra loro, per religione, costumi e colore della pelle: Erodoto descrive popolazioni diverse, e cosi' anche Ibn Battuta e Marco Polo; eppure non videro "razze" umane ma popolazioni. Poi quasi all'improvviso, il concetto di razza applicato all'uomo prende piede. E' possibile suggerire una spiegazione interessante di questo apparente paradosso della storia della cultura: Erodoto, come Ibn Battuta e Marco Polo, viaggiava prevalentemente a piedi, tutt'al piu' a cavallo; se usava una nave, era soltanto per la navigazione costiera, interrotta da frequenti approdi per il rifornimento. I navigatori del Rinascimento, da Colombo e Vasco Da Gama in poi, navigavano su navi a vela capaci di (rischiose) traversate oceaniche. In conseguenza della differenza nel modo di viaggiare, Erodoto vedeva gradienti genici e la transizione graduale di caratteri fisici da una popolazione ad un'altra; Colombo vide un brusco cambiamento dei caratteri fisici passando senza soste intermedie dalle popolazioni europee a quelle del nuovo mondo (discendenti probabilmente da popolazioni delle steppe asiatiche migrate attraverso lo stretto di Behring in epoca glaciale).
Origini remote della diversita' genetica umana
La speciazione umana e' avvenuta in Africa e la variabilita' genetica dell'uomo e' massima in Africa; piccoli gruppi umani sono migrati fuori dall'Africa in varie riprese, sfruttando periodi in cui le condizioni geoclimatiche erano favorevoli e si ritiene che l'umanita' extra-africana attuale sia il prodotto di un solo evento migratorio di un piccolo gruppo, avvenuto circa 100.000 anni fa. Eventi migratori precedenti, che avrebbero dato origine a popolazioni extra-africane piu' antiche, sono stati seguiti da estinzioni (ad es. per i Neanderthal) e solo una piccola parte dei genomi di queste popolazioni e' sopravvissuta fino a noi grazie a rari eventi di incroci con la popolazione extra-africana moderna.
La Tabella 5 ci mostra chiaramente che, anche se le popolazioni considerate si situano su un gradiente di variazione genica piu' o meno continuo, e' possibile stabilire un grado di differenziamento genico tra loro, con metodi piu' complessi ma concettualmente non dissimili da quelli utilizzati per la Tabella 2, ed e' evidente che gli inglesi sono geneticamente "piu' simili" agli italiani che ai giapponesi. Tabulando i gradi di differenza delle diverse popolazioni tra loro, e facendo ricorso agli opportuni strumenti matematici, e' possibile costruire grafi o alberi che rappresentino la distanza genetica tra le diverse popolazioni, e sembra ragionevole assumere che la distanza genetica rappresenti anche la distanza evoluzionistica da una originale popolazione comune. Le migrazioni avrebbero comportato, nel tempo, il differenziarsi delle varie popolazioni umane. Questo sarebbe stato anatema per i teorici del razzismo novecentesco, che pensavano che le razze fossero presenti fin da prima della speciazione umana.
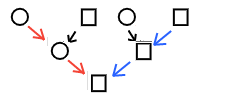
Poiche' ad ogni generazione si ha un rimescolamento di alleli paterni e materni, la struttura genetica delle popolazioni e' complessa, e la distinizione dalle popolazioni vicine e' in genere scarsa (cioe' le popolazioni non sono razze, e una etnia puo' includere piu' popolazioni). Ci sono pero' due sorgenti di DNA che non vanno incontro a rimescolamento e/ crossing-over e vengono quindi ereditate in forma immodificata: il DNA mitocondriale (mtDNA) ereditato per linea materna e la porzione non ricombinante del cromosoma Y, ereditato per via paterna. Lo studio di queste porzioni di DNA consente di ricostruire linee di discendenza che non sono genetiche, ma genealogiche: matrilineare e patrilineare. Si deve considerare che e' piu' frequente che una linea matrilineare o patrilineare si estingua (perche' non genera discendenti del sesso "giusto") piuttosto che nuove linee emergano, per mutazione. Queste analisi possono essere molto informative per ricostruire le origini di clan e popolazioni o le migrazioni umane di epoca preistorica, ma non si deve dimenticare che analizzano solo una piccolissima parte dell'informazione genetica complessiva; come si vede nella figura, vengono analizzate soltanto le linee rossa (matrilineare, del mtDNA) e blu (patrilineare, del cromosoma Y): l'analisi non considera le linee nere (di discendenza mista sia patrilineare che matrilineare). Di conseguenza lo stesso clan matrilineare potrebbe includere popolazioni geneticamente diverse, semplicemente perche' una donna ha contratto un matrimonio con un membro di un gruppo diverso da quello di origine, e trapiantato in quel gruppo la sua linea di discendenza matrilineare.