1) L'immunita' e' un complesso sistema di meccanismi fisiologici operanti a livello cellulare e umorale (cioe' di singole molecole proteiche in soluzione nei liquidi extracellulari) che ha la funzione di proteggerci dalle infezioni di parassiti, miceti, batteri e virus. Alcuni dei componenti dell'immunita' operano in modo aspecifico e sono attivi anche prima del contatto con l'agente infettante (immunita' innata), altri invece sono specifici e si sviluppano a seguito di un primo contatto con l'agente infettante (immunita' acquisita).
2) Fanno parte dell'immunita' innata: il lisozima, il complemento, i peptidi con attivita' battericida, gli interferoni, le citochine, e a livello cellulare, i granulociti e i macrofagi; fanno invece parte dell'immunita' acquisita gli anticorpi, e a livello cellulare i linfociti B e T.
3) Il lisozima e' una endoglicosidasi specifica prodotta dalle cellule delle mucose, specialmente quelle mucosa in contatto con l'ambiente esterno (ad es. nelle cavita' orale e nasale).
4) Il complemento e' un insieme di proteine presenti nel siero di sangue e prodotte dal fegato nella forma di precursori inattivi, che con un meccanismo a cascata proteolitica per molti versi simile a quello della coagulazione, va incontro ad un processo il cui esito finale e' l'attivazione di enzimi ad attivita' fosfolipasica capaci di danneggiare la membrana delle cellule procariotiche, causandone la morte. Fa parte del sistema dell'immunita' innata, ma puo' interagire con gli anticorpi che ne aumentano la specificita' e lo rendono in parte dipendente dall'immunita' acquisita. Le proteine del complemento sono termolabili e il complemento viene inattivato riscaldando il plasma a 56oC.
Le proteine del complemento sono individuate con la sigla C ed un numero. L'attivazione del complemento segue vari percorsi:
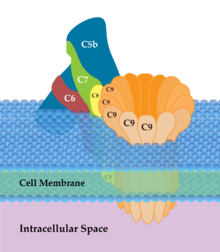
a) il percorso classico richiede l'attivazione del complesso C1, di solito mediata dall'interazione con un anticorpo. Il complesso C1 attivato attiva i componenti C2, C4 questi il componente C3. I componenti C2, C3 e C4 attivati si uniscono a formare un complesso multienzimatico (C5 convertasi) la cui funzione e' attivare il componente C5. Il componente C5 attivato recluta i componenti C6, C7, C8 e C9 che formano il complesso multienzimatico finale (complesso di attacco alla membrana) che si inserisce nello spessore del doppio strato fosfolipidico e ne degrada idroliticamente i componenti creando dei fori sulla membrana cellulare. Le cellule dell'organismo sono protette perché ospitano delle proteine di membrana capaci di inibire la funzione enzimatica del complesso di attacco alla membrana (CD35, CD46, CD55, CD59); i microorganismi patogeni in genere non possiedono nella loro membrana proteine capaci di inibire il complemento.
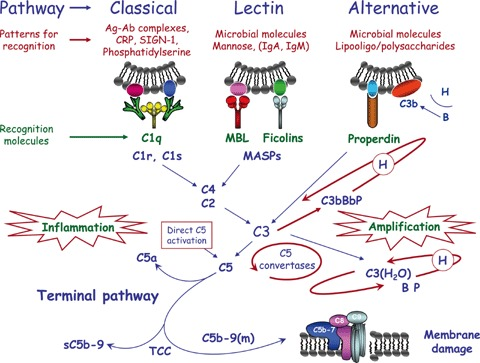
b) Il percorso alternativo non richiede l'intervento di anticorpi e sfrutta la proprieta' del C3 di autoattivarsi con basso livello di efficienza ed e' sempre attivo, indipendentemente dalla presenza di patogeni. Se non sono presenti microorganismi patogeni il C3 autoattivato viene degradato dai fattori circolanti H ed I; se invece sono presenti batteri, il C3 autoattivato aderisce alla loro parete e risulta protetto dall'inattivazione e si combina con il fattore P (properdina) che ne amplifica l'attivita'.
c) Esiste anche un terzo meccanismo di attivazione del complemento che interviene quando la parte batterica espone delle lectine (proteine capaci di combinarsi con disaccaridi e mucopolisaccaridi.
Il complesso finale, attivo, del complemento e' composto dai componenti C5, C6, C7, C8 e C9; questi ultimi formano la struttura tubulare che si inserisce nella membrana della cellula batterica e la perfora.
5) I polipeptidi antibatterici sono numerosi.
6) Gli interferoni sono proteine appartenenti a diverse classi (α; β; γ; λ) prodotte da varie cellule in risposta a infezioni virali, mutazioni cancerogene, etc. Di solito lo stimolo alla produzione di interferone e' la presenza nel citoplasma di un eccesso di RNA, in particolare di RNA a doppio filamento (caratteristico di molti virus). La cellula infettata da un virus attiva la biosintesi e la secrezione di interferone che va ad attivare le cellule vicine. L'effetto dell'interferone, tanto sulla cellula infetta che sulle cellule vicine, e' quello di promuovere la biosintesi di alcune proteine, tra le quali in particolare la RNAsi L e la proteina chinasi R (PKR). La PKR e' attivata dal RNA a doppio filamento (dsRNA) e fosforila specificamente la proteina eIF2, un fattore di attivazione della traduzione. L'eIF2 fosforilato perde l'attivita' biologica e questo riduce grandemente la biosintesi proteica nella cellula. La RNAsi L degrada l'RNA libero nel citoplasma, bloccando ulteriormente la sintesi proteica. Questo tipo di difesa e' suicida: la cellula infetta muore a causa del blocco della sintesi proteica. L'azione difensiva dell'interferone si esplica in questo: "suicidare" la cellula infetta prima del completamento del ciclo di replicazione virale. L'infezione virale di cellule attivate dall'interferone e' quindi abortiva e questo spegne la malattia.
| interferone | localizzazione del gene | cellula produttrice |
| IFN α (famiglia di circa 20 proteine diverse) | 9 | linfociti B e T |
| IFN β | 9 | vari tipi cell. spec. fibroblasti |
| IFN γ | 12 | linfociti T |
| IFN λ | 19 | vari tipi cell. |
7) Le citochine sono proteine che mediano la comunicazione tra cellule. Sotto stimolo di citochine una cellula puo' essere indotta a differenziarsi, replicarsi o morire. Svolgono varie funzioni nella regolazione del ricambio cellulare; inoltre, ai fini della difesa dalle infezioni virali, svolgono funzioni simili a quelle degli interferoni in quanto causano l'autodistruzione della cellula infetta prima che il ciclo virale abbia il tempo di completarsi. In effetti gli interferoni stessi possono essere considerati una specifica classe di citochine.
8) Gli anticorpi o immunoglobuline sono le proteine dell'immunita' specifica. Sono macromolecole costituite (in genere) da una coppia di catene polipeptidiche pesanti e una coppia di catene polipeptidiche leggere:
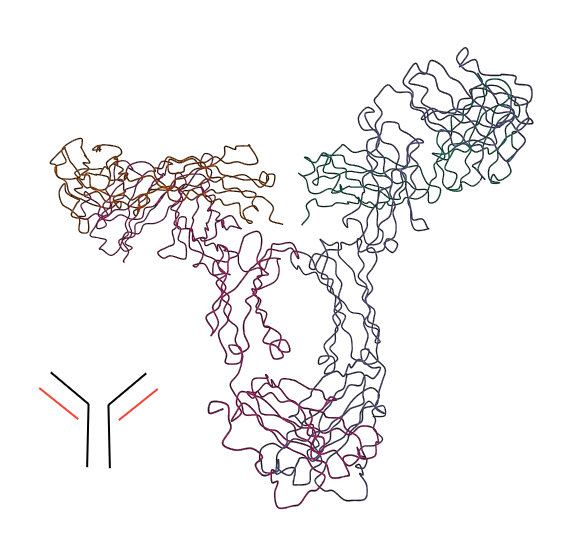
Le immunoglobuline appartengono a vari tipi; alcune hanno la struttura riportata in figura, altre sono multipli di quella (trimeri o pentameri).
Le parti fondamentali della molecola di immunoglobulina sono le tre estremita': le due estremita' simmetriche costituite da parti della catena pesante e leggera (antigen binding fragment, frammento AB, FAB) e l'estremita' costituita dalle due catene pesanti (frammento cristallizzabile, FC). I FAB si combinano con le molecole estranee all'organismo (in genere si tratta di molecole sulla superficie degli agenti infettanti, virus o batteri) e, sperabilmente, ne bloccano l'attivita' biologica; inoltre poiché l'Ig ha due FAB, collega tra loro multipli antigeni e ne causa la precipitazione, agglutinandoli in aggregati multimolecolari o multicellulari. Il FC attiva i macrofagi e promuove la fagocitosi dell'antigene.
La struttura delle immunoglobuline e' rigida, prevalentemente a foglietto γ.
Le Ig non sono tutte presenti alla nascita. Man mano che l'organismo incontra sostanze estranee (spesso prodotte da agenti infettanti quali virus, batteri e parassiti), i linfociti B vengono stimolati e vanno incontro a replicazione cellulare. Nella duplicazione del DNA le regioni terminali delle catene leggere e pesanti vanno incontro a ripetute mutazioni (regioni ipervariabili) e generano molte varianti clonali. I cloni di linfociti B che producono anticorpi capaci di combinarsi con l'antigene proliferano ulteriormente; gli altri vengono eliminati. L'immunita' dovuta agli anticorpi e' pertanto acquisita, anziche' innata. Se l'organismo riesce a produrre anticorpi efficaci in tempi brevi di solito guarisce dall'infezione. In un eventuale successivo incontro con lo stesso agente patogeno risulta immune perche' i cloni di linfociti B attivati rimangono latenti nelle stazioni linfatiche (cellule della memoria immunitaria) e possono essere riattivati molto rapidamente.
Nell'elettroforesi del siero le immunoglobuline costituiscono una frazione piuttosto grande (banda delle γ globuline) e sono le proteine che migrano piu' lentamente verso il polo positivo:
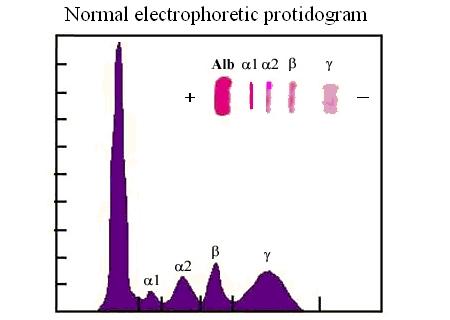
9) Self e non-self