1) La fosforilazione ossidativa ha lo scopo di convertire energia sotto forma di NADH in ATP. Questa funzione e' fondamentale perche' il ciclo di Krebs produce soprattutto NADH mentre la cellula necessita soprattutto di ATP. La fosforilazione ossidativa ha due componenti: la catena respiratoria e la ATPasi.
2) La catena respiratoria accoppia il trasferimento di elettroni dal NADH all'ossigeno (che viene convertito in H2O) allo spostamento di ioni idrogeno dalla matrice mitocondriale allo spazio intermembranario. I complessi multienzimatici della catena respiratoria risiedono nella membrana mitocondriale interna. E' interessante notare che nella respirazione cellulare si osserva la dissociazione tra il consumo dell'ossigeno e la produzione dell'anidride carbonica. La CO2 viene prodotta nel ciclo di Krebs; l'ossigeno viene utilizzato dalla catena respiratoria.
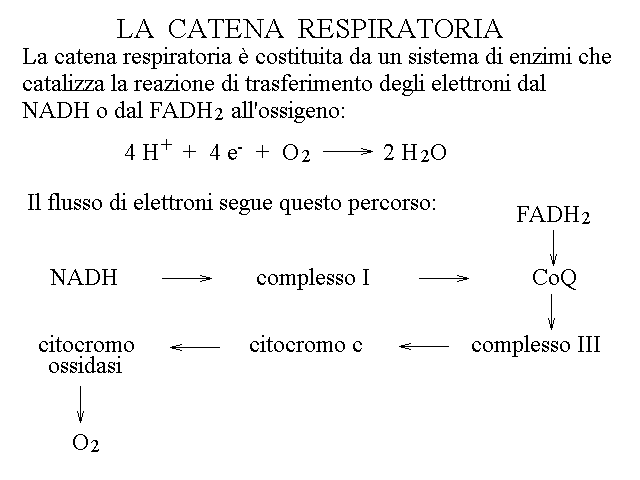 |
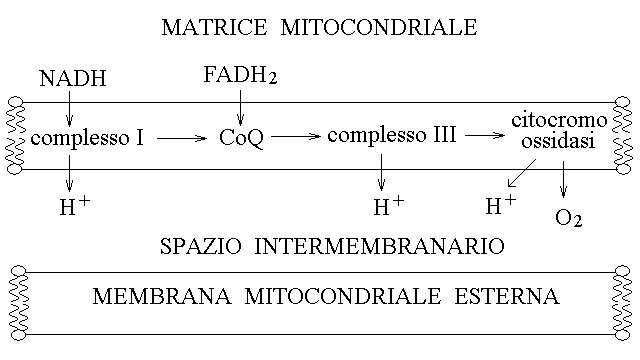 |
3) Protoni scalari e protoni vettoriali. Gli enzimi della catena respiratoria trasferiscono ioni idrogeno con due meccanismi distinti: (a) quando il trsaferimento di elettroni e' associato al legame o rilascio di ioni idrogeno questi vengono sempre prelevati dalla matrice e rilasciati nello spazio intermembranario (protoni scalari):
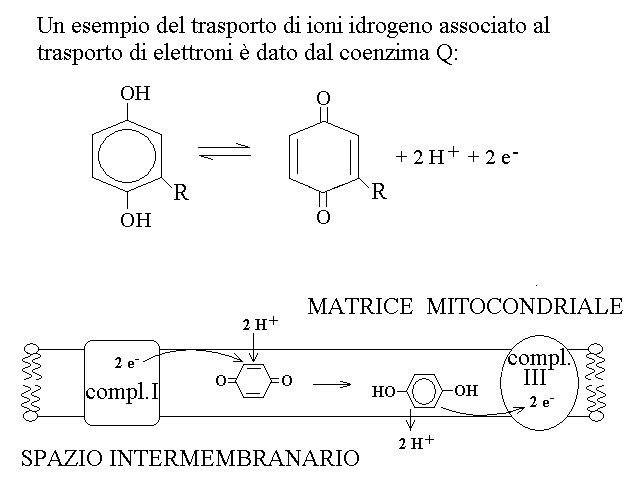
4) I complessi multienzimatici della catena respiratoria:
a) il complesso I accetta elettroni dal NADH e li trasferisce al Coenzima Q (ubichinone, un idrochinone sostituito). Trasferisce sia protoni scalari che vettoriali.
b) Il complesso II accetta elettroni dal FADH2 e li trasferisce al Coenzima Q. E' un enzima molto particolare che partecipa sia al ciclo di Krebs che alla catena respiratoria; nel ciclo di Krebs viene chiamato succinato deidrogenasi e ossida il'acido succinico a fumarico.
c) Il complesso III trasferisce elettroni dal Coenzima Q al citocromo-c; poiche' NADH, FADH2 e Coenzima Q accettano coppie di elettroni mentre il citocromo-c accetta singoli elettroni, ogni molecola di Coenzima Q ridotto riduce due molecole di citocromo-c.
d) Il citocromo-c e' una piccola emproteina nella quale il gruppo eme e' stabilmente esa-coordinato e passa da uno stato ossidato (in cui lo ione ferro ha carica +3) ad uno ridotto (in cui lo ione ferro ha carica +2).
e) La citocromo-c ossidasi (complesso IV) accetta 4 elettroni da 4 molecole di citocromo-c ridotto e li cede ad una molecola di ossigeno, formando 2 molecole d'acqua. Gli ioni idrogeno necessari sono prelevati dalla matrice. Inoltre la citocromo-c ossidasi trasferisce protoni vettoriali dalla matrice allo spazio intermembranario.
Una animazione del trasferimento degli elettroni attraverso gli enzimi e i coenzimi della catena respiratoria si puo' vedere in questo filmato realizzato dall'Universita' di Harvard.
5) L'ATPasi (ATP sintetasi). L'energia redox contenuta nel NADH viene convertita dalla catena respiratoria in energia chemioosmotica, cioe' legata alla creazione di un gradiente ionico tra i due versanti della membrana mitocondriale interna. Il gradiente viene sfruttato dall'ATPasi per la fosforilazione dell'ATP. L'ATPasi e' analoga ad una pompa di membrana che funziona pero' in modo inverso rispetto al solito: cioe' anziche' creare un gradiente con consumo di ATP, dissipa un gradiente con produzione di ATP:
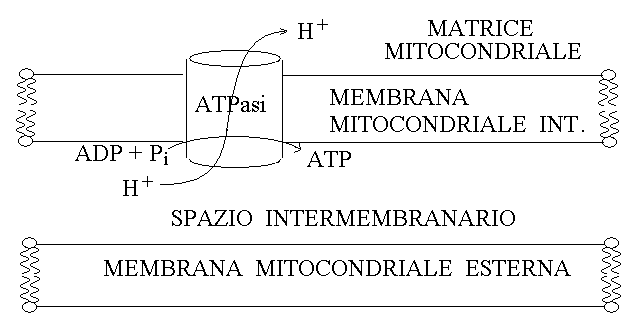
L'ATPasi e' una molecola complessa, formata da varie subunita' che compongono le sue parti intra- ed extra-membranaria; la reciproca rotazione di queste due parti e' responsabile del trasferimento degli ioni idrogeno e della fosforilazione dell'ADP come si puo' vedere in questo video realizzato dall'universita' di Harvard.
6) Equivalenze energetiche. Nelle condizioni piu' favorevoli, la riossidazione di una molecola di NADH trasferisce complessivamente circa 10 ioni idrogeno dalla matrice allo spazio intermembranario; quella di una molecola di FADH2 circa 6. L'ATPasi fosforila 1 ATP per ogni tre ioni idrogeno trasferiti dallo spazio intermembranario alla matrice; quindi si possono ottenere 3 ATP/NADH e 2 ATP/FADH2. In realta' questa equivalenza non viene di solito realizzata perche' la membrana mitocondriale interna non e' totalmente impermeabile ai protoni; in condizioni fisiologiche le equivalenze sono nell'ordine di circa 2,5 ATP/NADH e 1,5 ATP/FADH2. Alcuni ormoni regolano la permeabilita' della membrana mitocondriale interna ai protoni e disaccoppiano la fosforilazione ossidativa (cioe' disaccoppiano il trasferimento degli elettroni dalla fosforilazione dell'ATP); il principale e' la tiroxina.