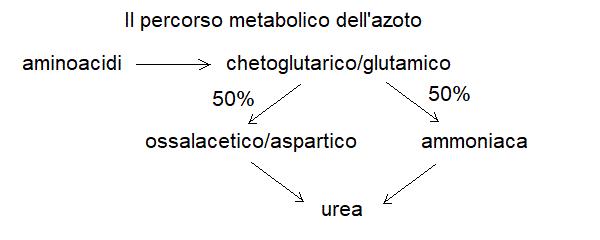
1) La dieta di qualunque animale include proteine, in genere in eccesso rispetto alle necessita' del ricambio; per questo parte degli aminoacidi devono essere utilizzati per produrre energia, sfruttando il ciclo di Krebs. Questo comporta due problemi:
a) gli aminoacidi devono essere deaminati e l'azoto deve essere eliminato
b) la matrice carboniosa dell'aminoacido (che dopo la deaminazione in genere si trasforma in un chetoacido) deve essere trasformata in acetil-CoA o in metaboliti della glicolisi.
Questa lezione descrive i processi metabolici di eliminazione dell'azoto; il metabolismo degli aminoacidi deaminati sara' descritto in lezioni successive.
2) La deaminazione degli aminoacidi produce ammoniaca (o acido aspartico, vedi oltre). L'ammoniaca e' tossica per il rene e puo' essere eliminata soltanto a concentrazioni molto basse. Gli animali che vivono in acqua possono produrre una urina diluita ed eliminare senza danno l'ammoniaca come tale: sono definiti ammoniotelici (il nome indica che l'ammoniaca e' per loro il prodotto terminale del metabolismo dell'azoto). Gli animali terrestri, allo scopo di risparmiare acqua, convertono l'ammoniaca in composti azotati meno tossici, che il rene possa concentrare nell'urina. Sono definiti ureotelici (se il metabolita terminale dell'azoto e' l'urea) o uricotelici (se il metabolita terminale dell'azoto e' l'acido urico). L'uomo, come tutti i mammiferi e' ureotelico.
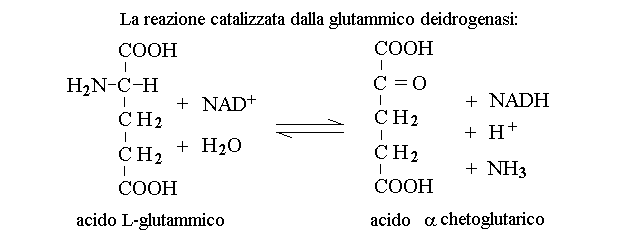
3) I processi di deaminazione degli aminoacidi e la produzione dell'ammoniaca. La maggioranza degli aminoacidi viene deaminata ossidativamente dalle transaminasi, che trasferiscono il gruppo aminico sull'acido α-chetoglutarico. I prodotti sono α-chetoacidi e acido glutamico. Le transaminasi utilizzano il PLP come coenzima, e sono enzimi citoplasmatici presenti soprattutto nelle cellule del fegato. La reazione catalizzata dalla glutamico-piruvico transaminasi (GPT; ALT), ad esempio, e' la seguente:
4) La produzione dell'urea avviene nel ciclo dell'urea che fu scoperto da Hans Krebs e Kurt Henseleit. La principale sede del ciclo sono gli epatociti; una piccola frazione dell'urea viene anche prodotta dalle cellule renali. Il ciclo converte una molecola di anidride carbonica, una di ammoniaca e una di acido aspartico in una molecola di urea e una di acido fumarico, secondo la seguente equazione generale:
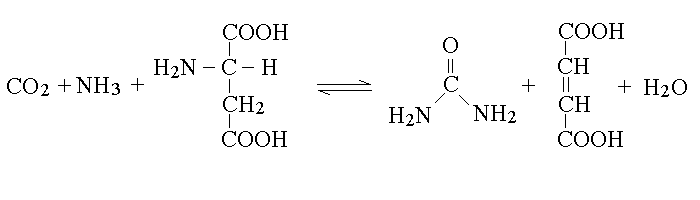
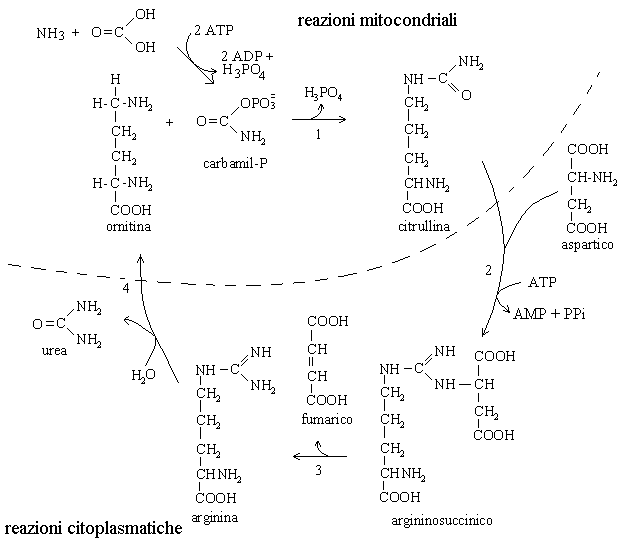
5) Gli scambi di metaboliti tra il citoplasma e il mitocondrio sono estremamente importanti, non solo per il ciclo dell'urea. Un elenco abbastanza aggiornato e completo si puo' trovare nell'appendice della pubblicazione The mechanism of transport by mitochondrial carriers based on analysis of symmetry di Robinson e co-autori (PNAS, USA, 2008, 105, 17766-).
6) Malattie del ciclo dell'urea: possono essere genetiche o acquisite; si manifestano con ridotta concentrazione serica dell'urea e iperammoniemia. Le forme genetiche sono rare, ma sono descritte per tutti gli enzimi del ciclo. Comportano iperammoniemia, convulsioni e coma; sono in genere letali nei primi anni di vita. Sono tutte ereditate come caratteri genetici autosomici e recessivi, con l'eccezione della carenza di ornitina transcarbamilasi, che e' X-linked. Si deve tenere presente che in alcuni casi il difetto genico comporta l'assenza totale o la mancata sintesi dell'enzima colpito; in altri casi, forse maggioritari, l'enzima mutante viene sintetizzato ed e' funzionale, ma e' instabile e si denatura rapidamente; pertanto la sua concentrazione nella cellula e' fortemente ridotta ma non e' zero. In aggiunta alle possibili mutazioni dei geni degli enzimi del ciclo si devono considerare quelle dell'antiporter ornitina/citrullina e quelle degli enzimi che sintetizzano l'ornitina a partire dall'acido glutamico.
Le iperammoniemie acquisite in genere si manifestano negli stadi terminali dell'insufficienza epatica cronica (epatiti croniche, cirrosi epatica, etc.). La prognosi e' infausta.
7) Regolazione del ciclo dell'urea. La carbamil-fosfato sintetasi e' regolata allostericamente dall'acetilglutamato, la cui sintesi a sua volta dipende dalla presenza di un eccesso di aminoacidi liberi nel citoplasma. L'ornitina transcarbamilasi e' un omotrimero, attivato dal carbamil-fosfato.
8) Biosintesi dei metaboliti necessari al ciclo. L'acido ossalacetico e' prodotto da varie vie metaboliche, la principale delle quali e' la carbossilazione del piruvato, operata dalla piruvato carbossilasi:
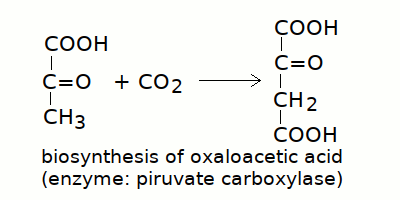
L'acido α-chetoglutarico e' prodotto nel ciclo di Krebs a partire dall'acido ossalacetico, oppure dall'acido glutamico e dalla glutamina presenti nella dieta.
L'ornitina e' un aminoacido non proteico, che e' richiesto in piccola quantita' nel ciclo e non e' fornito dalla dieta; puo' essere prodotto o a partire dall'arginina derivante dagli aminoacidi proteici, dall'enzima arginasi, oppure per transaminazione dalla glutamico semialdeide, a sua volta derivata dall'acido glutamico; questa via metabolica, che e' comune alla biosintesi di prolina e spermina puo' essere visualizzata a questo link.