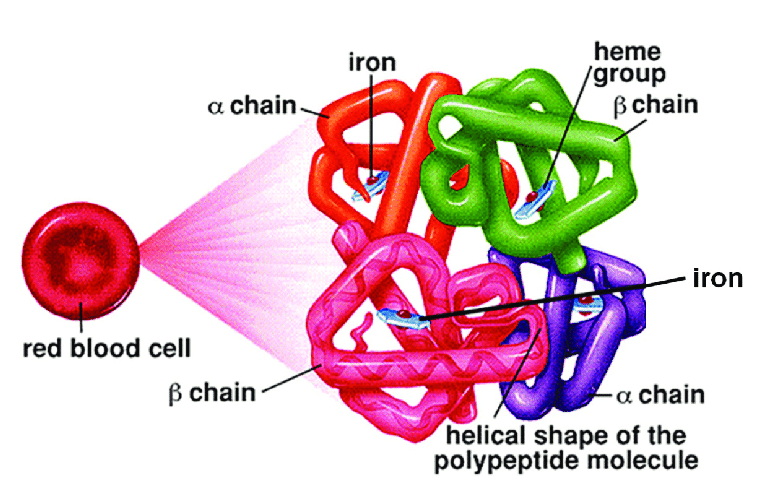
1) Come gia' spiegato nel Corso di Chimica e Propedeutica Biochimica, le proteine sono eteropolimeri lineari orientati di aminoacidi, legati con legami peptidici 2) Il polimero si ripiega su se stesso grazie agli angoli rotazionali fi e psi dei C alfa di ciascun aminoacido, formando strutture secondarie periodiche (alfa elica, foglietto beta) e non periodiche (loops, random coils) 3) Segmenti di struttura secondaria diversa possono a loro volta ripiegarsi su se stessi formando strutture globulari o fibrose (struttura terziaria); esistono inoltre proteine che hanno lunghi tratti di random coil disordinato (natively unfolded) 4) Le proteine possono associarsi fra loro e formare oligomeri composti da piu' catene polipeptidiche (struttura quaternaria); un esempio e' dato dall'emoglobina. La formazione di un oligomero composto da monomeri proteici richiede che esistano sui monomeri interfacce geometricamente e chimicamente complementari e sono possibili due casi: quello di interfacce eterologhe (cioe' diverse, come accade nel caso della tubulina), e quello di interfacce isologhe (come accade nel caso dell'emoglobina o di molti enzimi): 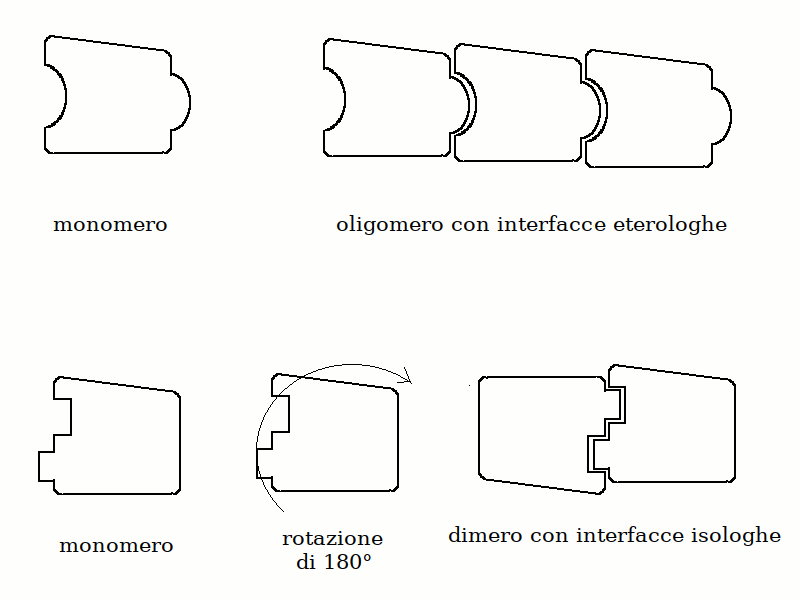 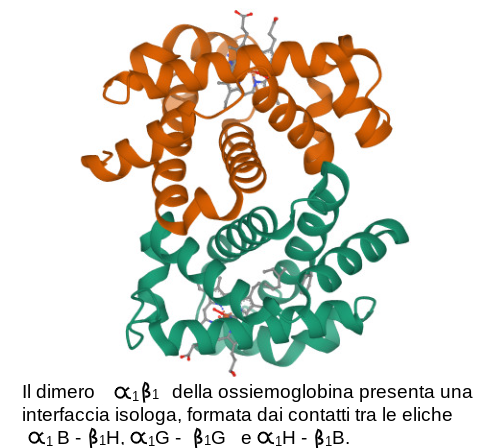 Una struttura particolarmente interessante e' quella delle proteine le cui subunita' si dispongono in modo da formare anelli; questi possono presentare sia interfacce eterologhe che interfacce isologhe. Un esempio e' dato dall'enzima purina nucleotide fosforilasi:  5) Le proteine natively folded sono strutture compatte nelle quali i residui aminoacidici sono praticamente in contatto tra loro e le cavita' hanno prevalentemente dimensioni subatomiche; le cavita' di dimensioni atomiche o superatomiche sono rare. L'interno delle proteine idrosolubili raccoglie in genere la maggioranza dei residui apolari (hydrophobic core). 6) Il ripiegamento di segmenti di struttura secondaria (ad es. alfa eliche) su se stessi comporta che i contatti tra residui aminoacidici nell'hydrophobic core possano verificarsi non solo tra residui che nella sequenza aminoacidica sono vicini o adiacenti, ma anche tra residui che nella sequenza sono molto lontani. Spesso le alfa eliche e i foglietti beta alternano residui apolari, rivolti verso l'hydrophobic core e residui polari rivolti verso la superficie della proteina. 7) Le proteine anche piu' compatte hanno moti vibrazionali interni a livello di singoli residui aminoacidici o di interi segmenti di struttura secondaria. Questi movimenti hanno grande importanza per la funzione proteica e dipendono dalla temperatura; pero' possono portare alla denaturazione della proteina stessa. 8) La struttura che la proteina assume all'interno della cellula e che e' capace di svolgere la funzione biologica si chiama nativa. In linea di massima la struttura nativa e' un minimo energetico e la proteina la raggiunge da sola, gia' durante la biosintesi. Ciononostante se esposta al calore, a solventi organici, a valori di pH molto alti o molto bassi, la proteina può perdere la struttura nativa (denaturazione) reversibilmente o irreversibilmente. 8) MODIFICAZIONI POST-TRADUZIONALI DELLE PROTEINE Molte proteine, soprattutto nelle cellule eucariotiche, subiscono modificazioni successive alla loro biosintesi. Queste possono essere reversibili o irreversibili e modificano l'attivita' biologica della proteina stessa; spesso la proteina neo-sintetizzata e' priva di attitvita' e la acquisisce solo in seguito alla modificazione post-traduzionale. Le modificazioni post-traduzionali sono frequenti nelle proteine extracellulari, piu' rare nelle proteine intracellulari. Le modificazioni post traduzionali possono avvenire sia all'interno della cellula che ha sintetizzato la proteina (spesso nel reticolo endoplasmatico o nell'apparato di Golgi) o all'esterno della cellula, nel liquido extracellulare, e richiedono enzimi specifici. Modificazioni post-traduzionali irreversibili - Taglio proteolitico limitato: la proteina neosintetizzata ha una catena polipeptidica piu' lunga della proteina attiva e la modificazione post-traduzionale consiste nell'idrolisi e rimozione di un frammento all'estremita' C- o N-terminale da parte di una proteasi specifica. Esempi sono: le proteine della coagulazione, il complemento, le proteasi digestive del pancreas, etc. Un diverso tipo di taglio proteolitico limitato si verifica nel caso in cui la proteina neosintetizzata debba essere tagliata in due frammenti che restano legati tra loro e che sono entrambi necessari per l'attivita' biologica; un esempio di questo caso e' dato dall'insulina. - Glicosilazione, con formazione di legami glicosidici tra residui di serina, treonina o tirosina e piccole catene oligosaccaridiche o singoli monosaccaridi. - Modificazioni selettive di singoli aminoacidi: ad es. gamma-carbossilazione degli acidi glutamici (nei fattori della coagulazione); idrossilazione di residui di prolina o lisina (nel collageno); idrossilazione di residui di Tyr (nelle amino-ossidasi a rame). - Aggiunta di cofattori legati covalentemente o di ioni metallici. Modificazioni post-traduzionali reversibili - Fosforilazione-defosforilazione di residui di Ser, Thr, Tyr, in molte chinasi ed altri enzimi intracellulari. - Acetilazione-deacetilazione di residui di Lys, ad es. negli istoni. - Formazione di ponti disolfuro tra residui di Cys (reversibili per riduzione; ad es. nella tioredossina). |
{kind=link}